|
| Eng |
Морской зверь Каспия «И водится там одна рыба, совершенно похожая на собаку,
с головой, ногами и хвостом.» В фауне Каспийского моря имеется единственное морское млекопитающее, принадлежащее к отряду ластоногих, каспийский тюлень. Его происхождение во внутриконтинентальном водоеме остается до сих пор дискуссионным. Существуют две основные гипотезы, объясняющие происхождение этого вида. Согласно одной из них, поддерживаемой зоогеографами, и среди них Л. С. Берг: тюлени заселили Каспийское море в четвертичный период, проникнув тем или иным путем с севера под влиянием ледника, оттеснившего на юг часть популяции предков близкородственного вида, кольчатой нерпы, которая проникнув в новый водоем, в конечном счете и преобразовалась в современного каспийского тюленя. По другой гипотезе, основывающей более древнее происхождение вида и базирующейся на палеонтологических данных, относящихся к миоцену и плиоцену, каспийский тюлень считается потомком тюленей подрода Pusa, обитавших в последовательно сменявшихся Сарматском, Меотическом, Понтическом и других бассейнах, предшественниках современного Каспийского моря. Ряд ископаемых остатков верхнетретичных и раннечетвертичных тюленей, найденных на периферии Каспия и в других южных районах, обнаруживает черты близкого родства с современным каспийским тюленем.
В палеоцене млекопитающие постоянно обитали на суше, но примерно 60 миллионов лет назад некоторые группы сухопутных четвероногих предпочли вернуться в море, породившее их, как и всех остальных животных на свете. Причиной этого попятного движения, вероятнее всего, был поиск пищи. Никто не знает чей предок первым ушел под воду, однако натуралист В. Шеффер в своей книге «Естественная история морских млекопитающих» попытался составить хронологическую таблицу или эволюционный календарь. Он полагал, что четырем группам морских млекопитающих соответствуют четыре временных отрезка, отделяющих их от наземных предков. Так, морской стаж калана ( отряд хищных), по Шефферу, исчисляется всего в 5 миллионов лет. Ластоногие (тюлени и моржи) появились по меньшей мере 30 миллионов лет назад. Возраст сирен (ламантины и дюгони) определяется минимум в 55 миллионов лет, китообразных (киты и дельфины) – в 60 миллионов лет. Морские млекопитающие, составляющие четыре интереснейшие группы класса Mammalia, насчитывают примерно 118 видов, которые ведут всецело или частично водный образ жизни, в своем питании полностью зависят от моря, и их строение приспособлено к жизни в морской среде - в частности, у них развились ласты, плавники или перепончатые конечности. В том, что они истинные млекопитающие сомневаться не приходится, ведь теплокровные обитатели моря обладают всеми характерными особенностями этого класса: четырехкамерное сердце, способность самок рожать живых детенышей и выкармливать их молоком. Зависимость ластоногих от суши достаточно убедительно свидетельствует, что их предки сменили наземную среду на водную позднее, чем сирены и китообразные. Имеющиеся в распоряжении ученых окаменелости ластоногих датируются сравнительно недавней эпохой миоцена (около 20 миллионов лет назад). Об их наземном происхождении убедительно говорит их анатомическое сходство с современными наземными млекопитающими: сохранившееся наружнее ухо (морские котики), носовая полость обычного строения и четко разделенные задние ласты, позволяющие, пусть и довольно неуклюже, передвигаться на суше. Pennipedia – в переводе с латинского «ластоногие» – очень верное определение, данное учеными этому отряду морских млекопитающих. Все ластоногие выходят на сушу или лед для рождения детенышей и линьки. Большинство из них спариваются на твердом субстрате, а некоторые предпочитают и спать на суше или льду. Две древние легенды, одна скандинавская, другая ирландская, легли в основу прелестной небывальщины, сочиненной В. Шеффером. «В преданиях людей, которые кормятся морем, часто говорится, что в темном омуте глаз иных тюленей кроются духи, взывающие к определнным людям. Наиболее восприимчивы к этому зову ирландцы, а также некоторые скандинавы, исстари живущие у моря. Эти тюлени, говорят они, на самом деле – рыбаки, так или иначе навлекшие на себя недовольство богов, которые и обрекли их вечно жить в волосатой шкуре и странствовать по воле ветра и волн. Когда же такому тюленю случится спасти тонущего моряка, он освобождается от звериного обличья, превращается в прекрасную деву и становится женой спасенного. Но детей от такого союза не бывает, и старые женщины в селении знают тому причину. Глаза у этих прекрасных созданий всегда темно-карие, тело – мягкое, и в те ночи, когда через окно устремляется внутрь сияние полной луны, они лежат в постели без сна. И ноги у них несколько холоднее, чем у обычных женщин.» Географическое распространение каспийского тюленя ограничено исключительно Каспийским морем, что подтверждает его автохтоннное происхождение. Животные встречаются по всему пространству моря, от прибрежных районов Северного Каспия до берегов Ирана. Их можно встретить, как в очень мелководных районах, так и в зоне больших глубин. В тоже время каспийский тюлень принадлежит к пагофильной (льдолюбивой) группе тюленей, т.к. биологически связан со льдами, на которых размножается и выкармливает детенышей, а также проводит большую часть периода линьки. На протяжении годового цикла каспийский тюлень для своей жизнедеятельности ни один раз прибегает к помощи твердого субстрата, кроме льда он использует для лежбищ различного рода острова, каменистые гряды. Побережий, заросших тростником и другими высшими растениями, он, как правило, избегает. С конца Х1Х века не утихает полемика о вреде, якобы наносимым тюленем рыбному промыслу на Каспийском бассейне. Астраханский рыбопромышленник М. Хлебцов в начале прошлого столетия даже приводил сведения о выедании тюленем белорыбицы из ставных сетей. Развёртывание килечного промысла в Южном Каспии в 50-х годах ХХ столетия породило новую волну разговоров среди промысловиков о конкуренции тюленя с промышленным рыболовством и, в частности, его отрицательном воздействии на лов кильки на электросвет. Выедание рыбы из сетей, питание обыкновенной килькой в закидном (морском) неводе у Дагестанского побережья и, наконец, концентрация тюленя у промысловых судов, ведущих промысел анчоусовидной кильки в Южном Каспии, лишний раз говорят о трофической пластичности ихтиофага, использующего так называемый «принцип накрытого стола». Тюлени питаются во все времена года, но с разной интенсивностью по сезонам - поздней осенью, зимой и ранней весной не столь интенсивно, как летом. В ледовый период животные, занятые размножением, и кормящие самки питаются менее регулярно и живут впроголодь, поэтому вряд ли можно считать преувеличением, что после зимней жизни впроголодь тюлени « набрасываются на пищу весной». Зимой в северной части моря тюлени едят бычков и мелких ракообразных: гаммарид, мизид и креветок. По обеспеченности пищей зимнее время – наиболее трудный период в жизни животных. Средний вес пищевого комка в этот период по данным КаспНИРХ составляет 19,8 г. Причем годовые вариации доминирования отдельных пищевых компонентов в питании тюленя, например, бычков или крабиков характеризует не абсолютный запас кормовых объектов, а районы залежек тюленя. По мере приближениия сроков щенки беременные самки, пользуясь любыми возможностями проникнуть вглубь ледового пространства, отыскивают наиболее прочные льды и , когда уже приходит пора щениться , вылезают на лед. Взрослые самцы до поры до времени остаются вместе с неполовозрелыми особями того и другого пола ближе к прикромочной полосе и начинают вылезать на льды несколько позднее, образуя отдельные, более тесные, сплоченные залежки. Их еще называют «косячным зверем». Зимой 1926 г. два российских исследователя С.Дорофеев и С.Фрейман, представляющих НИРХ (совр.ВНИРО) на тюленке «Михаил» в составе зверобойной экспедиции провели почти два месяца, с 23 января по 11 марта, во льдах Северного Каспия. Живя все это время вместе с тюленщиками, они имели возможность, одновременно с исследованиями биологии тюленя, непосредственно изучить во всех деталях технику промысла тюленя, быт зверобоев, ледовый режим Северного Каспия в районе работ промысловой экспедиции. Полученные ими данные по биологии каспийского тюленя до сих пор не потеряли своей актуальности. Вот как они описывают начало экспедиции: «Форт Александровский (совр.Баутино,Р.Казахстан.) 24.01. в 6 час. Пошла вся флотилия – 61 судно. Ветер SOO свежий. Через час подошли к поясинам льда, двигающимся по ветру. Обошли их, вышли опять на полосу чистой воды. 8 час. Позавтракали, впереди опять поясины. Будем пересекать в узком месте. Изредка отдельные тюлени…» В своих отчетах по окнчанию экспедиции авторы отмечали, что труд зверобоя на Каспии рисковый и опасный, без груза пришли 32% всех судов и еще 9% погибло. Деторождение или щенка у каспийского тюленя в массовом порядке происходит в конце января-начале февраля. Продолжительность беременности составляет 11 месяцев. Самка каспийского тюленя обычно приносит одного детеныша, редко двух. При рождении длина тела щенка составляет приблизительно 70-75 см, вес 3-4 кг. Подкожного жирового слоя новорожденный почти не имеет, волосяной покров белькового типа: густой, длинный, шелковистый, почти чисто белый, нередко с дымчато-сероватым налетом на спинной стороне. Период кормления длится около 4 недель. В первые дни после родов матуха почти не отлучается от щенка, кормя его многократно в разное время суток. В дальнейшем она может оставлять детеныша на более длительный срок, делая продолжительные перерывы в кормлении. На льду детеныш большую часть времени проводит во сне. Проснувшись, он начинает искать кормилицу и часто, не дождавшись ее, подползает к лазке. Через нее он заглядывает в воду, ожидая оттуда появление матери. Если ожидание становится долгим, он призывает ее голосом, похожий на плач ребенка. Для кормления детеныша матуха ложится на бок или даже почти на спину, открывая соски. Растет и развивается молодняк также быстро, как и у других тюленей. За короткий период лактации детеныш вырастает на 20% от своей первоначальной величины, достигая 85-90 см, а его вес увеличивается более чем в 4 раза, достигая 8-12 кг, и уже через 1,5-2 декады после рождения белек начинает линять. В начале линька едва заметна, но с каждым последующим днем его пушистая бельковая шерсть лезет все более и более интенсивно. Постэмбриональная линька продолжается около 2-х недель, и в это время детеныша называют «тулупкой». Полностью перелинявший детеныш напоминает копию взрослого тюленя и уже называется «сиварем». Лактация прекращается на последних стадиях линьки; но нередко матуха кормит еще и сиваря. Волосяной покров сиваря сверху почти однотонный, темно-серого цвета; пятна, если проступают, то не всегда ясны. От года к году, с каждой очередной линькой на спине все более заметно проступает пятнистый узор. Наиболее резко пятнистость проявляется у самцов. Линька у всех особей, начиная с годовалых, одна в году. Первыми начинают линять ощенившиеся самки, позже взрослые самцы, а затем неполовозрелые обоего пола. Далеко не все тюлени успевают перелинять на льдах, и линька продолжается на шалыгах и островах. Каспийский тюлень ежегодно с большой регулярностью предпринимает широкие перемещения. Весной, с исчезновением льда на севере Каспия, сосредоточенная там еще с конца осени – начала зимы масса тюленей, начинает постепенно отходить к югу. Процесс этот растянут на длительное время, поскольку движение к югу совмещается с усиленным питанием. К концу весны-началу лета большая часть поголовья тюленя перемещается в Средний и Южный Каспий. Животные мигрируют большей частью сильно разреженными и мелкими группами, не сбиваясь в тесные косяки. Двигаются они обычно не непрерывно в одном и том же направлении, в особенности в период миграции к югу, скорее смещаются на юг в процессе беспорядочных кочевок. Кормясь на пути, звери двигаются широким фронтом одни вблизи западных берегов моря, другие придерживаются восточных окраин, третьи – на большом расстоянии от берегов. Но все же тюлени ежегодно посещают одни и те же районы. Одной из причин ухода тюленей весной на юг из мелководной северной части моря считается сильное летнее прогревание воды, которого тюлени избегают. В средней и южной частях моря благодаря большим глубинам и постоянному перемешиванию водной толщи, перегревание поверхностных вод не происходит, а поэтому для тюленей создаются более благоприятные температурные условия. Вместе с тем немалое значение имеет и трофический фактор. Весь летний период до середины сентября тюлени находятся в воде, рассеянные в ней поодиночке или мелкими группами. Спать на воде они могут также хорошо, как и на суше. Активность их в это время определяется в основном ощущением голода: она продолжается с большей интенсивностью до полного насыщения. Лето и часть осени – это основная часть нагульного периода, в течение которого животные не только восстанавливают свои жировые запасы, но и накапливают его для предстоящей зимовки. Нагульный или морской период характеризуется интенсивным потреблением пищи и протекает в весенне-летнее время преимущественно в Среднем и Южном Каспии. Масса пищевых комков тюленя во время нагула в несколько раз превышают эти показатели для других сезонов, хотя кормовая база обеспечивается коротким рядом пищевых организмов, главным образом массовыми короткоцикличными видами рыб р.Clupeonella (кильками). Анчоусовидная тюлька в летний период, когда верхние слои воды слишком прогреты, и благоприятная для нее температура сохраняется в более тонком слое, держится на глубине 10-40 м, совершая незначительные суточные миграции. Это способствует образованию плотных скоплений кильки в небольшом по высоте слое воды, что создает комфортные условия тюленю для активного питания. Из других кормовых организмов, характерных для нагульного периода следует отметить атерину и бычков. Пищевой спектр тюленей, не покидающих летом пределов Северного Каспия и являющихся больными или ослабленными, поэтому выпадающими из трофических миграций, представлен бычками, воблой, молодью леща. Как правило они держатся вблизи островов. В российских водах такой рекреационной зоной для каспийского тюленя является о.Малый Жемчужный (рис.2). В феврале 2002 г. по решению Правительства РФ он объявлен Федеральным Памятником Природы. С приближением осени почти все откормившиеся тюлени начинают постепенно откочевывать обратно к северу. Их активность к этому времени резко снижается. Животные выползают на острова, покрывая их сплошной массой своих тел. Тюлени в это время спят на боку, нередко на спине или на животе. В ясные солнечные дни тюлени часто лежат на острове или на шалыге, в пасмурные, а особенно в дождливые дни они лежат неспокойно или совсем уходят в воду. Пролонгированием нагула являются предзимние концентрации тюленя в авандельте Волги и Урала. Осенний период характеризуется широким спектром пищевых объектов, что связано прежде всего с предзимними концентрациями частиковых рыб. Как показывают многолетние данные КаспНИРХ, в пищевом спектре «авандельтовых» зверей превалируют массовые стайные виды рыб (главным образом молодь): вобла, судак, густера, лещ. Для Урала ещё характерны сазан и жерех. Причем существует определённая избирательность по размерам жертв, рыб длиной более 25 см каспийский тюлень, как правило, избегает.
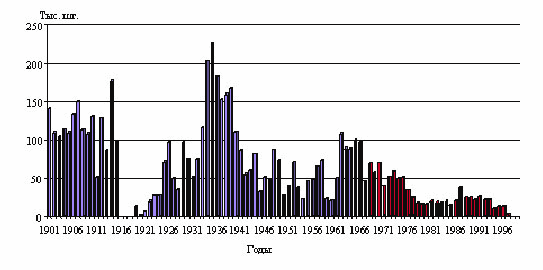
По типу питания каспийский тюлень ближе всего стоит к хищникам- ихтиофагам, т.к. доля других кормовых организмов (ракообразных) в его пищевом рационе незначительна, менее 1%. Элементы плотоядности, приписываемые отдельными авторами каспийскому тюленю из-за нахождения в его желудке перьев водоплавающих птиц скорее всего являются биологическим нонсенсом. Все посторонние предметы, относимые к категории балластного корма (перья птиц, кожура картофеля и др.) захватываются тюленем случайно и в основном молодыми особями. Глубина, на которую могут погружаться каспийские тюлени в поисках пищи по данным известного специалиста по каспийскому тюленю Б.И. Бадамшина составляет 85м. С приближением зимы и первым появлением льда лежбища на шалыгах и островах распадаются, и вся, подошедшая масса тюленей размещается на большей площади и держится преимущественно у кромки льда. Таким образом заканчивается годовой цикл жизни морского млекопитающего на Каспии. Каспийский тюлень имеет длительную промысловую историю. Достаточно сказать, что в официальный реестр промысловых зверей каспийские тюлени попали во времена правления русской царицы Анны Иоанновны, в 1740 г., а с 1867 г. КаспНИРХ владеет полной статистикой промысла. Размах колебаний добычи морзверя за прошедшее время достаточно велик: от нескольких тысяч до сотен тысяч голов (рис 3). К более-менее цивилизованному промыслу тюленя на Каспии удалось прийти только в последней трети ХХ столетия с введением жестких сроков и лимитирования добычи. Одновременно были прикрыты те разновидности тюленьего промысла, которые явно вредили запасам каспийского тюленя. Особенно сильный пресс промысла испытывала популяция каспийского тюленя в 30-е годы прошлого столетия, когда в течение пяти лет добывалось в среднем до 165 тысяч голов в год. В результате численность каспийского тюленя сократилась почти в двое с миллиона особей до 500-600 тысяч голов. Дальнейшее сокращение сырьевых ресурсов популяции на 20 % мы уже связываем с загрязнением экосистемы Каспийского моря. Кризис воспроизводства для каспийского тюленя прежде всего выразился в сокращении маточного стада и увеличении яловости потенциальных производителей, половозрелых самок.
«Кумулятивный политоксикоз» - такой диагноз для популяции каспийского тюленя был поставлен на состоявшемся в 1992 г. в Астрахани Международном Симпозиуме по проблемам экологии и болезней диких и сельскохозяйственных животных. Природа продолжает нам преподносить сюрпризы. С 1997 по 2000 г.г. на Каспии наблюдалось несколько крупномасштабных выбросов тюленя. В апреле- июне 2000 г. произошла самая большая гибель каспийского тюленя за прошедшее столетие. Погибшие животные регистрировались сначала на островных залежках и морской акватории Северного Каспия, а затем в западной части Среднего и Южного Каспия. В общей сложности на островах Северного Каспия и береговой полосе Казахстана, РФ (Дагестан) и Азербайджана гибель тюленя по нашим данным и сведениям местных природоохранных организаций составила порядка 25-30 тысяч особей. По факту гибели тюленя было осуществлено несколько экспедиций в северные районы гибели тюленя: на Зюйд-вестовую шалыгу (Казахстан) и о. Малый Жемчужный (РФ). Как показали проведенные исследования, патологоанатомическая картина и клинические признаки обследованных животных с островных залежек, удаленных друг от друга на расстояние 320 км, были примерно одинаковыми. В результате иммунологического анализа у 75% животных были выявлены патологические изменения в иммунной системе. Бактериологический анализ показал 100%-е инфицирование паренхиматозных органов (печени, почек, селезенки) и крови тюленей разнообразной микрофлорой. Исследования с помощью “Тест-системы иммуноферментной для диагностики чумы плотоядных”, предоставленной нами новосибирскими учеными из ГНЦ ВБ «Вектор», дали положительные результаты на присутствие вируса чумы плотоядных в популяции каспийского тюленя. Токсикологический анализ показал в свою очередь, что обнаруженные количества ХОП, тяжелых металлов и нефтяных ароматических углеводородов не подтверждают версию химического отравления с летальным исходом, но могут свидетельствовать о хроническом токсикозе животных, который способствовал ослаблению иммунитета и, как следствие, провоцированию инфекционных заболеваний. Наиболее вероятной причиной токсикоза является кумулятивное действие нефтяного и пестицидного загрязнений. По ряду признаков (быстрое, но не мгновенное распространение болезни, клинические симптомы) можно было предположить, что популяция каспийского тюленя поражена инфекционной эпизоотией. По последним данным института диагностировано заболевание тюленей сложной комбинированной инфекцией: пастереллезом и сальмонеллезом в сочетании с чумой плотоядных. Основной причиной элиминации тюленей является чума плотоядных, а бактериальные заболевания (пастереллез, сальмонеллез) являются секундарными инфекциями. Проведенные нами исследования нашли подтверждение в материалах английских, азербайджанских и казахских ученых. Как уже было сказано выше, развитие инфекционных заболеваний тюленя на Каспии в 2000 г. было обусловлено подавлением иммунной системы животных вследствие загрязнения экосистемы моря. Толчком же к развитию заболеваний могли послужить крайне неблагоприятные зимние условия. Чрезвычайно мягкая зима со среднемесячной температурой, в феврале на 7-90 превышающей норму, не могла не отразиться на ледовых процессах в Северном Каспии. Слабый ледовый покров и сокращенный ледовый период нарушили экологические характеристики существования вида. Линька животных происходила не на ледовых залежках, а в условиях большей скученности на шалыгах, расположенных вдоль восточного побережья, периодическое затопление которых под влиянием нагонных ветров усугубляло состояние линяющих тюленей. Высокая концентрация тюленей на островах привела к неудовлетворительному развитию и истощению животных, способствовала развитию инфекций. По последним данным в «рекреационных» зонах каспийского тюленя в Северном Каспии (о. М. Жемчужный) продолжает циркулировать вирус чумы плотоядных, поэтому существует опасность рецидива эпизоотии.
Кроме химического загрязнения, как оказалось, в последние годы не меньшую опасность для морских экосистем представляет биологическое загрязнение. Собственно говоря, антропогенное вселение чужеродных видов организмов в Каспий началось в начале ХХ столетия. С развитием судоходства и рыбохозяйственной деятельности во второй половине прошлого века этот процесс усилился. Вселение чужеродных организмов, так называемых ксенобионтов, в замкнутый водоем со слабой экологической валентностью автохтонных видов и низким биоразнообразием трофических уровней не может не нанести ущерба экосистеме. Появление же хищного гребневика мнениопсиса, который совсем недавно сотрясал экосистему Азово-Черноморского бассейна, может привести к наиболее драматическим изменениям в экосистеме Каспийского моря. По определению специалистов, как быстроразмножающийсяся, самооплодотворяющий гермофродит, новый вселенец является идеальным объектом для колонизации водоема. Даже в водоемах естественного ареала гребневик в условиях обильной кормовой базы способен быстро формировать огромную численность и биомассу, достаточные для истребления большей части продукции зоопланктона, а также существенно влиять на сообщества мезо- и ихтиопланктона. Каспийский тюлень нагуливается в летний период в Среднем и Южном Каспии, где его ареал соприкасается с формирующимся ареалом мнениопсиса. По наблюдениям ихтиологов в последние 2-3 года на Каспии под влиянием теплых зим происходит перегрев в летнее время поверхности моря, что негативно отражается на скоплениях анчоусовидной тюльки. Кроме того, устойчивая высокая поверхностная температура вызывает повышенное испарение, обуславлившее увеличение осолонения вод. Климатические изменения, по-видимому, оказались позитивным фактором для вселенца в силу его высокой экологической и биологической валентности. Численность гребневика резко повысилась, в результате чего он заполнил весь фотический слой пелагиали Южного Каспия и проник в среднюю часть моря. Появление нового, сильного конкурента планктоноядных рыб привело к резкому сокращению кормовой базы каспийских килек в 2001 г. в Южном Каспии в 10 раз, в Среднем – в 6 раз и, как следствие, к снижению численности анчоусовидной тюльки, являющейся одним из основных объектов питания каспийского тюленя в нагульный период. Ухудшение трофических условий не могло не сказаться на морфофункциональном состоянии постнагульных животных. В 2001 г. яловость самок достигла 79,8%, что превысило показатели ранее наблюдавшихся кризисных ситуаций в 1989-1990 и 1995 гг. По данным экологического мониторинга линейно- весовые показатели яловых самок были ниже среднемноголетних: масса тела – 41,7 кг, толщина жира – 4,9 см, масса хоровины – 21,2 кг, или 51% от массы тела. Средняя масса тела всех обследованных животных составила 41 кг, что ниже прошлогодних данных на 10%. Таким образом, можно сделать вывод, что снижение линейно-весовых показателей и накопления жировых запасов и, как следствие, уменьшение репродуктивного потенциала популяции связаны, по-видимому, с нарушением пелагических трофических связей в биоценозе моря из-за вселения мнемиопсиса. Популяция каспийского тюленя находится в неразрывной связи с другими обитателями Каспийского моря и всей экосистемой в целом. Об этом мы должны помнить всегда. Поэтому в условиях нарастания техногенного воздействия на экосистему моря в связи с разведкой и предстоящей разработкой углеводородного сырья мы не должны снижать мониторинговый контроль за эндемиком, т.е. видом который нигде больше не встречается. Л.Хураськин, рук. группы маммологов,
|   |
||||
| © 2000-2026 гг. Центр охраны дикой природы. Все права защищены |