|
| Eng |
К.А. Роговин Большой тушканчикAllactaga major (Kerr, 1792)
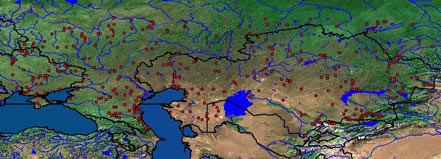
Подвиды: Allactaga major major Kerr 1792, (syn. chachlovy, decumanus, fuscus, intermedius, macrotis, vexillarius ) – Украина, Европейская часть России, Приуралье, Западный, Центральный и Восточный Казахстан, юг Западной Сибири (на восток до долины р. Иртыш); A. major spiculum (Lichtenstein, 1825 (syn. – brachyotis ) – Западная Сибирь от Иртыша до Оби; A. major djetysuensis (Shenbrot, 1991) – Южный и Юго-восточный Казахстан и Северная Киргизия (Семиречье). Представитель подрода Allactaga s. str., куда входит наряду с Allactaga elater, A. severtzovi, A. vinogradovi и, вероятно, A. hotsoni (Шенброт, 1984а; Шенброт и др., 1995). По строению черепа, коренных зубов и наружных мужских гениталий наиболее примитивный представитель подрода. Реальные филогенетические связи не прослежены. По-видимому, состоит в отдаленном родстве с нижнеплиоценовым Allactaga (Paralactaga) varians из Восточного Казахстана (Шенброт, 1984). Географическая изменчивость хорошо выражена по окраске, размерам и пропорциям тела и черепа, по строению наружных мужских гениталий. В качестве самостоятельных подвидов различными авторами были признаны в разное время A. m. spiculum (Lichtenstein 1825), A. m. decumanus (Lichtenstein 1825), A. m. vexillarius (Eversmann 1840), A. m. chachlovi (Martino 1922), A. m. fuscus (Ognev 1924), A. m. intermedius (Ognev 1948). Б.С. Виноградов (1937) отрицал существование подвидовой дифференциации большого тушканчика. Напротив, С.И. Огнев (1948) признавал существование описанных выше подвидов за исключением A. m. chachlovi (последнего рассматривал в составе A. m. vexillarius) Последняя ревизия подвидовой систематики по большому набору признаков, проведенная Г.И. Шенбротом (1991), показала существование трех хорошо выраженных подвидов. Обычный, широко распространенный вид, населяющий степи и полупустыни Евразии. Ареал вида имеет облик сплошного массива и одного изолята (Семиреченский участок – Юго-восточный Казахстан, Киргизия). Площадь изолята около 82,4 тыс. км 2 . Существование связи с основным ареалом, однако, нельзя исключить полностью (Шенброт и др., 1995). Дальше других видов проникает на север, заходя за 56° с.ш. В северной части ареала при сохранении общих его очертаний во второй половине 20 века отмечалось локальное вымирание вида в результате распашки целинных участков степи и сокращения площадей пастбищ и выгонов. Так, вплоть до середины 90-х годов встречался в Московской области (окрестности Серпухова). Ныне исчез там из-за уничтожения пригодных местообитаний (данные О.Н. Шекаровой). Размеры наиболее крупные среди тушканчиков вообще. Имеет облик типичного пятипалого тушканчика: сравнительно короткое тело (187-230 мм), хвост в 1,3 раза длиннее тела (250-305 мм), короткие передние и длинные задние конечности (длина ступни около 45% длины тела; 85-101 мм). У подвидов A. m. major и A. m. djetysuensis средняя длина тела составляет соответственно 205 и 215 мм, длина хвоста – 279 и 273 мм, ступни – 93 и 92мм, высота уха 57 и 59 мм, масса тела 319 г и 301 г. Две особи подвида A. m. spiculum из коллекций имели одинаковую массу тела равную 335 г (Шенброт и др., 1995). Боковые пальцы задних лап короткие, далеко не достигают своими концами до основания средних; внутренний палец немного длиннее наружного. Коническая мозоль в основании пальцев ступни крупная, высокая с широким основанием. Голова округлой формы с хорошо выраженным шейным перехватом. Мордочка слегка вытянутая, но широкая, с хорошо выраженным пятачком. Уши сравнительно короткие (высота их у разных подвидов от 23 до 29% длины тела). Окраска верха спины широко варьирует географически от интенсивной буровато-охристой или буровато-серой до бледной серовато-желтой (песчаной). Темная продольная струйчатость сравнительно хорошо выражена в задней части спины. Задняя часть верха головы имеет сходную со спиной окраску, но несколько более светлую. Окраска постепенно светлеет к концу мордочки, но вокруг носа обычно хорошо развито черное кольцо. Наружные уши покрыты короткими буровато-серыми волосками; вдоль наружного края уха и его вершины – узкая белая кайма. Щеки светлые, почти белые. Бока тела светлее спины за счет осветления оснований волос. Наружные поверхности бедер окрашены сходно со спиной. По бедру проходит короткая поперечная полоса белого цвета. Губы, горло, грудь, брюшко, предплечья и кисти, а также внутренние поверхности бедер чисто белые. Плюсна сверху покрыта короткими белыми волосками, а сбоку немного более длинными бурыми или черными волосками. Снизу ступня голая. Боковые поверхности пальцев задних конечностей несут буровато-белые удлиненные, плотно прилегающие к поверхности пальцев волоски, не образующие щетки. Стержень хвоста сверху ярко окрашен, в целом сходно со спиной. Хвостовое знамя великолепно развито, имеет форму птичьего пера. Сверху оно двухцветное. Черное поле большое, начинается еще до знамени на стержне хвоста. Белое поле на конце знамени также хорошо развито; его длина с учетом индивидуальной и географической изменчивости, в общем, близка к длине черного поля (сопоставима с его длиной). Снизу черное поле знамени имеет форму ласточкиного хвоста лишь на границе с белым полем. На другом конце его черная окраска сравнительно далеко распространяется вверх по стержню хвоста. Череп мощного сложения с широко расставленными скуловыми дугами. Кондилобазальная длина черепа в выборках из разных частей ареала, включая разные подвиды, изменяется от 40,1 до 46 мм, при близких средних значениях у разных форм от 42,4 до 43,7 мм. Скуловая ширина изменяется в пределах ареала от 35,5 до 36,2 мм при средних значениях у разных форм от 31,9 до 33,8 мм. Вертикальная ветвь скуловой дуги одинаковая на всем протяжении и по ширине примерно равна горизонтальной ветви. Рострум широкий, умеренной длины. Носовые кости сильно расширяются в передней части. Ширина костного неба на уровне вторых верхних коренных превышает ширину этих зубов в 1,5-1,8 раза. Диаметр верхнего предкоренного зуба в 1,7-1,9 раза меньше длины третьего верхнего коренного. Коренные зубы мезодонтные с низкими коронками и террассированной жевательной поверхностью (Шенброт и др., 1995). Glans penis имеет сердцевидную форму с вытянутой и притупленной на конце вершиной и слегка уплощен в дорзовентральном направлении. Поверхность его покрыта одновершинными направленными назад шипиками. Шипики увеличиваются в размерах от вершины к основанию. На латеральных лопастях шипики объединены в 6-8 правильных косых рядов по 10-15 в каждом (Шенброт, 1995). Хромосом в диплоидном наборе 48. Число плеч аутосом 92 (Воронцов и др., 1971). Половой диморфизм по окраске, размерам тела и черепа не выражен. В поле по пропорциям внешнего строения, великолепно развитому знамени безошибочно определяется как один из представителей пятипалых тушканчиков рода Allactaga . От других крупных представителей Allactaga , встречающихся совместно ( A. sibirica, A. severtzovi ) отличается крупными размерами. Отловленные экземпляры большого тушканчика в большинстве случаев отличаются на глаз от предыдущих видов по длине ступни и по отсутствию белого кольца перед черным полем знамени. Большие тушканчики не совершают резких прыжков при беге и уходят от преследования равномерными плавными, но мощными прыжками. Создается впечатление, что тушканчик летит над землей. В отличие от A. sibirica относительно чаще использует защитные норы.
Биотопы Один из наиболее экологически пластичных видов тушканчиков. Населяет разнообразные местообитания. На севере ареала основным лимитирующим фактором служит густота травостоя. Поселения приурочены к открытым участкам с разреженной растительностью. В степной зоне часто селится по обочинам грунтовых дорог, по кромкам полей, на выгонах, по пологим склонам балок и оврагов (Огнев, 1948; Попов, 1960, Токарский, Ронкин, 1990). В степях Казахстана и Западной Сибири населяют низкотравные луга с солонцеватыми почвами, берега соленых озер и степных речек, опушки сосновых боров, обочины дорог, полей, выпасы (Серебренников, 1929; Белов, 1931; Строганова, 1954; Глотов, Телегин, 1969 и др.). В пустыне и пустыне большой тушканчик может быть встречен практически во всех типах местообитаний, за исключением слабо закрепленных и подвижных песков. Предпочитают участки с легкими суглинистыми почвами и с мозаикой растительности из полыней и суккулентов. Привязанность к таким богатым кормами участкам усиливается к югу (Фенюк, 1928; Белослюдов, 1948; Ходашова, 1960; Сабилаев, 1967а; Лобачев и др., 1976а). В Семиречье большой тушканчик населяет широкий спектр местообитаний, поднимаясь в горы до 1650 м над уровнем моря. Наиболее характерны для него подгорные суглинистые участки полупустыни с полынями и низкорослыми злаками (Касаткин, 1958; Мазин, 1973; Катаевский, 1976).
Обычный, но не достигающий высокой численности вид. Численность (как общая численность, так и локальная плотность) увеличивается с севера на юг. В степях на европейской части ареала плотность зверьков по оценкам разных авторов изменяется в пределах от 0,5 до 5 особей/га (Андриевский, Елисеев, 1952; Калабухов, 1959; Миронов и др., 1965; Сурвилло и др., 1985; Магомедов, 1987, наши данные (Калмыкия, Дагестан). В Западной Сибири плотность населения составляет 0,1-0,2 особи/га (Никифоров, 1959). В степных районах Северного Казахстана плотность достигает 2-3, реже 4-6 особей/га (цит. по Шенброт и др., 1995). В различных пустынных районах Западного Казахстана отмечалась плотность 0,2-0,8 особей/га, а также численность от 0,5 до 6 особей на 10 км автомобильного ночного маршрута (цит. по Шенброт и др., 1995). В Семиречье сравнительно многочисленен – в среднем 6-7, до 14 зверьков на 10 км ночного автомобильного учета (Катаевский, 1976). В Алма-Атинской области плотность может достигать 8 особей/га, численность 3 особей на 1 км ночного автомобильного учета (Мазин, 1973). Резких колебаний численности год от года, по-видимому, не бывает. По типу питания большой тушканчик относится к всеядным грызунам. В пищевом рационе примерно в равной мере могут участвовать семена, зеленые, подземные части растений и корма животного происхождения. Данные выборочных оценок разных авторов о соотношении типов корма в диете различаются как вследствие географической изменчивости питания, так ив результате значительной сезонной изменчивости. Большой тушканчик легко переходит с одного типа корма на другой, в зависимости от его обилия в данном месте и в данный сезон. Обязательно во всех случаях присутствие калорийного корма. Это могут быть насекомые, семена, луковицы, которые тушканчики ловко выкапывают. Так в Северном Приаралье в составе содержимого желудков по объему летом насекомые составляли 8%, семена – 35% подземные части растений – 42% и зеленые части растений 15% (Шенброт, 1980). В Дагестане во все периоды активной жизни тушканчиков зеленые и подземные части растений присутствовали в равном количестве (Магомедов, Субботин, 1985). В Северном Приаралье в первой половине лета насекомые составляли 27% состава диеты, семена – 15%, подземные части растений – 37%, зеленые части растений 21% (данные Г.И. Шенброта из Шенброт и др., 1995). В Алма-Атинской области Казахстана весной большие тушканчики питались в основном насекомыми и подземными частями растений, летом – семенами (плодами), а осенью – зелеными частями растений и насекомыми (Мазин, 1973). Строго зимоспящий вид. Залегают в спячку осенью с наступлением постоянных заморозков. Обычно это происходит в сентябре, реже в октябре. Пробуждение наступает в марте-апреле (Фенюк, 1928, 1929; Бёме, 1952; Катаевский, 1976). Продолжительность спячки в разных регионах от 4 месяцев (Хохлов, 1985) до 6-6,5 месяцев (Фенюк, 1928, 1929; Бёме, 19 5 2, Катаевский, 1976). Перед залеганием сильно жиреют. Масса тела может увеличиваться почти вдвое. Запасы полостного жира достигают 22,5 г (Фенюк, 1929; Сабилаев, 1968б, данные Г.И. Шенброта из Шенброт и др., 1995). У большого тушканчика существует три типа нор: летние постоянные, зимовочные и защитные. Постоянная нора начинается плотно забитым землей начальным ходом, расположенным почти горизонтально под поверхностью земли. Длина этого хода может достигать 6 м. От горизонтального хода в не забитой его части (примерно от его середины) отходит круто наклонный ход, который ведет к гнездовой камере. Этот ход часто еще и поворачивает под углом 90°. Основной горизонтальный ход (продолжение начального хода), открывается другим своим концом наружу. Используемое входное отверстие норы обычно забито земляной пробочкой. От основного горизонтального хода могут отходить отнорки немного недоведенные до поверхности земли и служащие запасными выходами (обычно один, редко – 2-4). Гнездовая камера всегда присутствует в выводковых норах самок. В норах других зверьков ее может не быть. Гнездо устроено из сухих стеблей трав, кусочков мха, шерсти пуха и перьев и имеет форму шара, заполняя собой камеру диаметром 13-19 см. Вход в нору округлый диаметром 5,5 см. Общая длина основных ходов составляет 125-615 см, глубина залегания гнездовой камеры – 40-110 см. (Фенюк, 1928, 1929; Колесников, 1934; Мазин, 1973; данные Г.И. Шенброта). Зимовочные норы, по-видимому, представляют собой перестроенные летние постоянные норы, однако восстановить их первоначальную структуру не удавалось, поскольку горизонтальный ход в описанных норах отсутствует. Обычно состоят из одного равномерно углубляющегося хода. Ход круто поворачивает в начале под углом 90°, затем идет более или менее прямо наклонно. В норе имеются две гнездовые камеры на разных уровнях – одна к конце хода, другая в его средней части на конце небольшого отнорка на глубине 45-70 см. Длина ходов зимовочных нор 310-500 см. Максимальная глубина 155-250 см (Фенюк, 1929; Попов, 1960; Мазин, 1973) . Защитные норы имеют простое строение и состоят из одного прямого или слегка изогнутого, равномерно углубляющегося хода. Длина его 80-200 см, а глубина 50-80 см. Вход в нору хорошо заметен, имеет форму эллипса (диаметры его 10-13 см х 7-9,5) и характерный подход к норе в виде желоба (Фенюк, 1928, 1929; Шенброт и др., 1995). Большой тушканчик – вид со строго ночной активностью. Лишь расселяющиеся молодые в редких случаях встречались среди дня (Фенюк, 1928). Тушканчики, как правило, выходят на поверхность через 40-45 минут после восхода солнца, заканчивается активность за 30-50 минут перед заходом солнца (Фенюк, 1928). Время выхода зависит от сезона и упитанности зверьков. В Западном Казахстане весной можно встретить большого тушканчика сразу после захода солнца. В сравнении с мелкими видами пятипалых тушканчиков A. major более осторожен при выходе из норы. Прежде чем покинуть нору, отодвинув мордочкой земляную пробку, он долго остается в норе, высунув лишь голову, прислушиваясь. Выйдя из норы, тушканчики широко перемещаются по участку обитания, делая кратковременные остановки для кормления. Большую часть времени на поиск корма и питание большой тушканчик тратит лишь во второй половине ночи (Магомедов, 1989). Судя по результатам учетов зверьков на маршрутах, пик активности наблюдается в первой половине ночи (Андриевский, Елисеев, 1952; наши данные). Это может быть связано, не с количеством зверьков на поверхности, а с их высокой двигательной активностью в начале ночи. На участке обитания большого тушканчика всегда присутствуют 2-3 защитные норы, которые зверьки посещают во время ночных прогулок. Одно из объяснений тяготения большого тушканчика к защитным норам (как и других крупных видов Allactaga ) в отличие от малого тушканчика состоит в меньшей маневренности крупного и стремительно передвигающегося грызуна в случае атаки со стороны хищника (в том числе при нападении с воздуха). В спокойном состоянии большой тушканчик передвигается на двух ногах шагом или рысью, при быстром движении используют асинхронный рикошет (последовательные толчки сначала одной, затем другой лапой). При нормальном беге длина прыжка составляет 80-125 см (данные Г.И. Шенброта). Максимальная скорость бега – 40-50 км/час (Огнев, 1948), или м/сек (Роговин, 1999). При преследовании спасаются бегством в одном направлении на расстояние 100м и более, часто в направлении к защитной норе. Участки обитания специально не изучались. Их радиус по наблюдениям за отдельными экземплярами составлял 140-190 м. (Роговин, 1999). В природе редко контактируют друг с другом, избегая взаимодействий. В неволе взрослые особи агрессивны. Характерен один сильно растянутый период размножения. В разных частях ареала беременные самки встречаются с марта до июля (в основном апрель-июнь). За сезон размножения некоторые самки успевают принести по два выводка. Известны случаи совмещения беременности с лактацией (Мазин, 1975; Лобачев, Шенброт, 1976). Молодые в год рождения в размножении не участвуют. В выводке от 1 до 8, обычно 3-6 детенышей (Фенюк, 1928, 1 9 29; Колесников, 1934; Сорокина и др., 1985; Лобачев, Шенброт, 1976; Сабилаев, 1968; Мазин, 1975; Катаевский, 1976). Беременность по косвенным оценкам длится около 25 дней. Детеныши появляются на свет голыми и слепыми с пропорциями тела новорожденного квадрупедального грызуна. Лактация и совместное пребывание детенышей с самкой не менее полутора месяцев. Минимальная масса тела вышедших на поверхность молодых (120-125 г). При этом самка еще продолжает кормить детенышей молоком. Молодые переходят к самостоятельной жизни при достижении массы тела 180-220 г (Мазин, 1975). Максимальная продолжительность жизни в природе не менее трех лет (Лобачев, Шенброт, 1976). Линька в юго-восточном участке ареала протекает с мая по сентябрь. Пик приходится на июль-август (Катаевский 1976; Слудский и др., 1977). Андриевский И.В., Елисеев Л.Н. 1952. Материалы по экологии тушканчиков в районе государственной полезащитной лесной полосы Камышин – Сталинград // Бюлл. МОИП, отд. биол. Т. 57, вып 6. С. 19- 24. Белов В.Н. 1931. Обзор грызунов Северного Казахстана // Тр. по защите растений Сибири. Т. 1(8). С. 164-182. Белослюдов В.А. 1948. Млекопитающие пустыни Бетпак-Дала // Изв. АН Каз. ССР. Т. 63, сер. зоол., вып. 8. С. 51-78. Бёме Л.Б. 1952. К познанию грызунов Казахской складчатой страны // Бюлл. МОИП, отд. биол. Т. 57, вып. 2. С. 45-53. Бiльский Б. 1929 Поширення великого земляного зайця ( Allactaga jaculus Pall. = A. saliens Gm.) на Правобережнiй Украiнi // Тр. Фiз.-Мат. Вiд. Всеукр. АН. Т. 13, вып. 1. С. 147-180. Виноградов Б.С. 1937. Фауна СССР. Млекопитающие. Т. 3, вып. 4. Тушканчики. М.-Л.: Изд. АН СССР. 197 с. Воронцов Н.Н., Малыгина Н.А., Раджабли С.И. 1971. Хромосомы тушканчиков (Rodentia, Dipodidae) // Зоол. журн. Т. 50, вып. 12. С. 1853-1860. Глотов И.Н., Телегин В.И. 1969. Распространение большого тушканчика и алтайского цокора // Биологическое районирование Новосибирской области. Новосибирск: Наука. С. 59-62. Катаевский В.Н. 1976. К экологии большого тушканчика Таласской долины // Тр. Кирг. ун-та. Сер. биол. н. Вып. 15/2. С. 82-86. Калабухов Н.И. 1959. Особенности размещения грызунов и методика учета их численности в полосе ильменей на правобережье Волги // География населения наземных животных и методы его изучения. М.: изд. АН СССР. С. 280-286. Касаткина Н.В. 1958. Материалы по распространению грызунов в предгорьях Заилийского Ала-Тау // Тр. Среднеаз. н.-и. противочумн. ин-та. Алма-Ата. Т. 4. С. 218-221. Колесников И.И. 1934. Вредные грызуны каучуконоса тау-сагыз. М.-Ташкент: Среднеазиатск. отд. ОГИЗа. 96 с. Лобачев В.С., Шенброт Г.И. 1976. Особенности размножения тушканчиков // Бюлл. МОИП, отд. биол. Т. 81, вып. 4. С. 38-52 Лобачев В.С., Сабилаев А.С., Шенброт Г.И. 1976. Распределение по биотопам и численность тушканчиков Приаралья // Фауна и экология грызунов, вып. 13. М.: Изд-во МГУ. С. 13-40. Магомедов М.-Р.Д. 1987. Половая структура популяции некоторых зимоспящих видов грызунов // Экология. Вып. 2. С. 13-20. Магомедов М.-Р.Д. 1989. Зависимость питания и состояния популяций тушканчиков от динамики кормовых ресурсов // Журн. общ. биол. Т. 50, вып. 6. С. 824-835. Магомедов М.-Р.Д., Субботин А.Е. 1985. Изучение интенсивности питания тушканчиков // Зоол. журн., Т. 64, вып. 5. С. 731-737 Мазин В.Н. 1973. Тушканчики юго-востока Казахстана и их хозяйственное значение: Автореф. дисс. канд. биол. наук. Алма-Ата. 26 с. Мазин В.Н. 1975. Материалы по размножению тушканчиков рода Allactaga на юго-востоке Казахстана // Экология. Вып. 4. С. 53-60. Миронов Н.П., Карпузиди К.С., Клименко И.З., Колесников И.М., Лисицын А.А., Нельзина Е.Н., Ширанович П.И., Ширяев Д.Т., Яковлев М.Г. 1965. Источники и переносчики чумы и туляремии. М.: Медицина. 195 с. Никифоров Л.П. 1959. Опыт биосъёмки населения млекопитающих Тоболо-Ишимской лесостепи // География населения наземных животных и методы его изучения. М.: Изд. АН СССР, С. 7-23. Огнев С.И. 1948. Звери СССР и прилежащих стран. Т. 6. М.-Л.: Изд-во АН СССР. 559 c. Попов В.А. 1960. Млекопитающие Волжско-Камского края. Казань: Изд-во АН СССР. 468 с. Роговин К.А. 1999. К вопросу о происхождении бипедальной локомоции у грызунов (Экологические корреляты двуногого бега тушканчикообразных) // Зоологический журнал 78 (2): 228-239 Сабилаев, А.С. 1967. Размещение и численность тушканчиков в Каракалпакии и прилежащей территории // Экология млекопитающих и птиц. М.: Наука. С. 60-66. Сабилаев А.С. 1968. Материалы по распространению и экологии большого тушканчика на Каракалпакском Устюрте // Узб. биол. журн. Вып. 1. С. 57-59. Серебренников М.К. 1929. Материалы по систематике и экологии грызунов Южного Зауралья // Ежегодник зоол. музея АН. Т. 29, вып. 2. С. 251-284 Слудский А.А., Бекенов А., Борисенко В.А., Грачев Ю.А., Исмагилов М.И., Капитонов В.И., Страутман Е.И., Федосенко А.К., Шубин И.Г. 1977. Млекопитающие Казахстана. Т. 1, ч. 2. Алма-Ата: Наука. 536 с. Сорокина З.С., Сорокина З.Н., Потапова Е.А., Газиев С.Г.-М., Кочетов В.А., Мялковский В.А., Глушко Н.В. 1985. Тушканчики восточного Предкавказья // Тушканчики фауны СССР. М: Всесоюз. териол. о-во. С. 66-67. Строганова А.С. 1954. Млекопитающие степного и полупустынного Заволжья // Тр. зоол. ин-та АН СССР. Т. 16. С. 30-116. Сурвилло А.В., Попов Н.В., Денисов П.С., Денисова Н.Г. 1985. Распространение, численность и эктопаразиты тушканчиков Калмыцкой АССР // Тушканчики фауны СССР. М: Всесоюз. териол. о-во. С. 70-71. Токарский В.А., Ронкин В.И. 1990. Большой тушканчик в Харьковской области // Вестн. Харьк. ун-та. Т. 346. С. 86-87. Фенюк Б.К. 1928. К биологии тушканчиков // Мат-лы к познанию фауны Нижнего Поволжья. Вып. 2. С. 1-43. Фенюк Б.К. 1929. Еще о биологии тушканчиков и о мерах борьбы с ними // Мат-лы к познанию фауны Ниж. Поволжья. Вып. 3. С. 1-52. Ходашева К.С. 1960. Природная среда и животный мир глинистых полупустынь Заволжья. М.: изд. АН СССР. 131 с. Хохлов А.Н., Бичерев А.П., Сигида С.И. 1985. К экологии большого тушканчика на Ставрополье // Тушканчики фауны СССР. М: Всесоюз. териол. о-во. С. 185-186. Шенброт Г.И. 1980. Сравнительная экология тушканчиков пустынь Турана. Автореф . дисс. канд. биол. наук. М.: МГУ. 1980. 22 с. Шенброт Г.И. 1984. Строение коренных зубов и филогения пятипалых тушканчиков подсемейства Allactaginae (Rodentia, Dipodidae) // Сб. тр. зоол. музея МГУ. Т. 22. С. 61-92. Шенброт Г.И. 1991. Ревизия подвидовой систематики пятипалых тушканчиков рода Allactaga фауны СССР // Тр. зоол. ин-та АН СССР, Т. 243. С. 42-58. Шенброт Г.И., Соколов В.Е., Гептнер В.Г., Ковальская Ю.Г., 1995. Млекопитающие России и сопредельных регионов. Тушканчикообразные. М.: Наука. С 1-573.
|   |
||||||||||||||||||||||||
| © 2000-2026 гг. Центр охраны дикой природы. Все права защищены |