|
| Eng |
К.А. Роговин Малый тушканчикAllactaga elater (Lichtenstein, 1825)
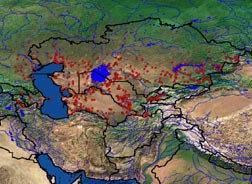
Подвиды: Allactaga elater elater (Lichtenstein, 1825) (syn. – kizliaricus, strandi, heptneri) – Северо - восточное Предкавказье , большая часть Казахстана кроме Восточного ( исключая юго - восток Прибалхашья , Алакольскую и Зайсанскую котловины ); A. elater caucasicus (Nehring, 1990) – Айзербайджан , крайний восток Грузии ; A. elater dzungariae (Thomas, 1912) – Юго - Восточное Прибалхашье , Алакольская котловина , юг Китайской Джунгарии ; A. elater zaisanicus (Shenbrot, 1991) – Зайсанская котловина , вероятно север Китайской Джунгарии ; Allactaga elater indica (Gray, 1842) – Юго - Восточный Туркменистан , Афганистан , Пакистан , Восточный Иран ; A. elater aralychensis (Satunin, 1991) – Армения , Нахичевань , Северо - Восточная Турция , Северо - Западный Иран ; A. elater turkmeni Goodwin, 1940 – Центральный и Северный Иран , возможен в Юго - Западном Туркменистане. Представитель подрода Allactaga s. str ., куда входит наряду с Allactaga major, A. severtzovi, A. vinogradovi и, вероятно, A. hotsoni (Шенброт, 1984; Шенброт и др., 1995). По строению черепа, коренных зубов и наружных мужских гениталий занимает промежуточное положение между A. vinogradovi и A. hotsoni . Реальные филогенетические связи не прослежены. Возможно, происходит от Protalactaga sp . из Северного Причерноморья (Шенброт и др., 1995). Географическая изменчивость хорошо выражена по окраске, размерам и пропорциям тела и черепа, по строению наружных мужских гениталий. В качестве самостоятельных подвидов различными авторами были признаны в разное время A. e. elater (Lichtenstein, 1895), A. e. indica (Gray, 1842), A. e. caucasicus (Nehring, 1900), A. e. aralychensis (Satunin, 1901), A. e. kizljaricus (Satunin, 1907), A. e. dzungariae (Thomas 1912), A. e. strandi (Heptner, 1934), A. e. turkmeni (Goodwin, 1940), A. e. heptneri (Pavlenko, Denisenko , 1976). Б.С. Виноградов (Виноградов, 1937; Виноградов Аргиропуло, 1941) считал реально существующими лишь три подвида – A. e. elater, A. e. indica (в современной трактовке) и A. e. vinogradovi (ныне признаваемый отдельным видом, Шенброт, 1991). С.И. Огнев (1948), напротив, признавал подвидовую самостоятельность всех известных к тому времени форм. Последняя ревизия подвидовой систематики по большому набору признаков, проведенная Г.И. Шенбротом (1991), показала существование семи хорошо выраженных подвидов, распадающихся по признакам строения наружных гениталий на две группы. Шесть из них обитают на территории бывшего СССР, присутствие седьмого возможно (см. выше). На большей части сплошного ареала обычный, и даже многочисленный вид. Сравнительно малочисленны в настоящее время популяции номинативного подвида из Северо-западного Прикаспия, изолированные от основного ареала рекой Волгой (Шенброт и др., 1995), (площадь изолированной части ареала около 92800 км 2 ). В связи с быстро развивающимся остепнением и сокращением площади местообитаний пустынного типа численность малого тушканчика в этом регионе имеет тенденцию к сокращению (наши данные по Калмыкии 1997-2000 г.). В Средней Азии в 70-80е годы 20 века ареалы A. elater elater , а также A. elater indica (в Туркменистане) заметно сократились из-за распашки больших массивов целинных земель. В настоящее время сведения о состоянии популяции A. e. indica полностью отсутствуют. A. elater dzungariae на востоке основного ареала вида – обычный представитель локальных сообществ грызунов. Подвиды Закавказья, A. elater caucasicus и A. elater aralychensis , имеют небольшие изолированные ареалы площадью около 52000 км 2 и 12000 км 2 соответственно. Локальная численность этих подвидов также, по-видимому, не бывает высокой (Эйгелис, 1980; Рабинович, Ахундов, 1975). Другой, изолированный от основного ареала подвид, – A. elater zaisanicus , локально обычен, но имеет ограниченный ареал площадью около 30400 км 2. Самый мелкий представитель пятипалых тушканчиков, типичного для рода Allactaga облика. Тело сравнительно короткое, хвост в 1,5-1,6 раза длиннее тела с хорошо выраженным, напоминающим перо птицы концевым "знаменем". Длина тела взрослых от 90 мм до 120 мм (в среднем для выборок из разных географических районов – 105-112 мм). Длина хвоста от 140 до 190 мм (в среднем от 160 до 173 мм). Средняя длина ступни изменяется в выборках из разных регионов от 49 мм до 5 5 мм (пределы: 43-58 мм). Ступня удлиненная; ее длина составляет 44-50% длины тела. Боковые пальцы задних конечностей далеко не достигают своими концами основания средних. Внутренний палец немного длиннее наружного. Мозоль в основании пальцев ступни крупная, высокая с закругленной вершиной. Уши умеренно длинные, около 29-33% длины тела. Длина наружного уха варьирует от 30 мм до 37 мм (в среднем – от 35 до 38 мм). Масса тела варьирует в пределах 32-77 г. В разных частях ареала средняя масса тела в выборках составляет 45-58 г (Шенброт и др., 1995). Основная окраска верха головы и спины глинисто-серая с примесью песчано-желтых или охристых, иногда розоватых тонов и темной размытой продольной струйчатостью. В некоторых популяциях по спине вдоль хребта проходит неясная темная сероватая полоса с размытыми краями. Бока тела, передняя часть головы окрашены светлее спины. Граница окраски спины и боков резко обозначена за счет того, что основания волос на боках не серые, а чисто белого цвета. Наружные поверхности бедер в верхней половине окрашены сходно со спиной. По середине их проходит широкая поперечная полоса белого цвета, снизу окаймленная темно-серым. Нижняя часть бедер ярко охристая. Эта окраска распространяется на наружную поверхность голени. Губы, горло, грудь, брюшко и передние части щек чисто белые. За ушами небольшие белые пятна. Плюсна и пальцы сверху и сбоку покрыты белыми волосками, снизу они голые. Плюсна и пальцы по краям оторочены узкими полосками охристых с черными вершинами волосков по 2-3 мм. Стержень хвоста сверху окрашен светлее и ярче спины. Бока и низ хвоста значительно светлее верха. Хвостовое знамя трехцветное сверху, белое или охристое в основании (5-15 мм), черное в средней части (20-35 мм) и чисто белое на конце (15-20 мм длиной). Снизу черное поле бывает разорвано продольной белой полоской вдоль стержня. В других случаях белый цвет более или менее глубоко вклинивается спереди и сзади в срединное черное поле, имеющее форму ласточкиного хвоста. Наибольшая длина вибрисс 65-80 мм. Голова округлая, мордочка укороченная, широкая, с хорошо выраженным пятачком. Череп облегченного строения. Кондилобазальная длина черепа варьирует от 24,0 до 27,7 мм в среднем 25,3-26,6 мм в выборках из разных частей ареала. Вертикальная ветвь скуловой дуги в своей нижней половине по ширине сходна с горизонтальной ветвью, а в верхнем отделе резко расширена. Верхние резцы очень слабо выступают вперед и направлены почти строго вниз. Межглазничный промежуток широкий, в среднем 8,5-9,6 мм. Слуховые барабаны сравнительно некрупные, но заметно вздутые. Ширина костного неба на уровне вторых верхних коренных превышает ширину этих зубов в 1,5-2 раза. Верхний предкоренной зуб по своему диаметру в 1,4-1,8 раза меньше длины третьего верхнего коренного. Коренные зубы мезодонтные, но с низкими коронками (Шенброт т др., 1995). Хромосом в диплоидном наборе 48. Число плеч аутосом – 92. (Воронцов и др., 1971). Кариотип идентичен таковому у A. vinogradovi. Половой диморфизм по окраске, размерам тела и черепа не выражен (Шенброт и др., 199 5). В поле от сходных по размерам и обитающих совместно тарбаганчика ( Pygeretmus pumilio ) и тушканчика Житкова ( P. zhitkovi ) отличается более длинными ушами, никогда не имеющем утолщения хвостом и хорошо развитым концевым "знаменем". При отгибании вперед конец наружного уха доходит или выступает за конец мордочки. От близкого Allactaga vinogradovi в полевых условиях по внешним признакам (без анализа черепа и гениталий) не отличим. От всех вышеназванных видов хорошо отличается защитным поведением. В случае преследования спасается бегством с резкими прыжками из стороны в сторону. В норы не заходит.
Малый тушканчик населяет разнообразные местообитания в полупустынной и пустынной зонах. Тип субстрата и проективное покрытие многолетней растительности служат ведущими факторами выбора местообитаний видом (Шенброт и др., 1995). Предпочитает супесчаные и легкие суглинистые почвы, однако, встречается и на плотных глинистых и щебнистых субстратах, и на участках уплотненных засоленных песков. В настоящих песках не живет, но нередко в них забегает. Обитатель разнообразных ассоциаций растительности с участием кустарников и полукустарничков. Избегает лишь больших полностью лишенных растительности пространств (обширные такыры) и густо заросших участков. Оптимальными для вида служат участки пустыни, где пятна многолетней растительности из небольших кустов и высоких трав перемежаются с полностью лишенными растительности пространствами (Шенброт и др., 1995). В Закавказье обитает в равнинных и подгорных солянковых полупустынях с глинистыми и щебнистыми поверхностями. Избегает опесчаненных участков с густой растительностью из полыней и злаков (Аргиропуло, 1939; Эйгелис, 1980). В Казахстане и Средней Азии наиболее многочисленен в полынно-солянковых и полынно-эфемеровых пустынях близ границ песчаных массивов, такыров, солончаков (Исмагилов, 1961; Ходашова, 1953; Ефимов, Зархидзе, 1964; Сабилаев, 1967; Лобачев и др. 1976; Шенброт, 1981; и др.). На большей части ареала обычный, а на юге Казахстана и в Средней Азии местами многочисленный вид. Номинативный подвид, A. elater elater в основном малочисленен на северо-западе ареала (Северо-западный Прикаспий), но даже здесь местами обычен. В дельте р. Терек отмечалась плотность в 6-9 особей/га (Магомедов, 1987, 1989; наши данные). В Калмыкии плотность достигает 4-6 особей/га (наши данные). В центральной части ареала (Западный и Центральный Казахстан – Средняя Азия) прослеживается тенденция к увеличению численности с севера на юг. Если близ северной границы ареала в Западном Казахстане описаны плотности в 0, 1 особь/га (Миронов и др., 1965), в 0,2-1 особь/га (Самарин и др., 1985) и численность, не превышавшая 8-9 особей (в среднем 3-4 особи) на 10 км автомобильного ночного маршрута (Алтухов и др., 1969), то на полуострове Мангышлак и на Устюрте отмечалась численность в 10-35, до 50 особей на 10 км ночного автомобильного учета (Ефимов, Зархидзе, 1964; Лобачев и др., 1976; Слудский и др., 1977; и др.). Еще южнее в Кызылкумах на территории Узбекистана известны плотности в 2-11 особей/га, 1-7 особей/га (Андрушко, 1939; Шенброт, 1981); в Южных Кызылкумах после весеннего размножения на участке эфемерово-полынной подгорной пустыни нами была отмечена плотность около 40 особей/га. На востоке ареала отмечалась как высокая, так и низкая численность вида (Шенброт и др., 1995). Наибольшие показатели известны для Южного Прибалхашья и Алакольской котловины ( A. elater djungariae ): обычно 4-20, в отдельных местах до 80 особей на 10 км ночного автомобильного учета, или не редко 10-20, в исключительном случае до 40 особей/га (Мазин, 1973; Кузнецов и др. 1985 и др., Шенброт и др., 1995). В Зайсанской котловине на 10 км автомобильного маршрута встречается в среднем 0,4, на отдельных участках до 12 особей A. elater zaisanicus на 10 км автомобильного ночного маршрута (Слудский и др., 1977; Прокопов, 1985). Для подвида A. elater indica из юго-восточной Туркмении известна плотность до 8-10 особей/га (Нургельдыев, 1960; Шенброт и др., 1995). В Восточном Закавказье малый тушканчик, A. elater caucasicus , видимо, немногосисленен (Эйгелис, 1980). Однако оценки затруднены из-за отсутствия сопоставимых данных. Плотность A. elater aralychensis в Карабахской долине на отдельных участках достигает 11 особей/га (Рабинович, Ахундов, 1975). Межрегиональные различия в численности малого тушканчика могут быть связаны с существенными перепадами численности в разные годы. Для дельты Терека (Северо-западный Прикаспий) Г.И. Шенброт приводит данные, свидетельствующие о 9 кратных изменениях численности за ряд лет. (Шенброт и др. , 1995). Он же свидетельствует о 8-кратном изменении численности в течение трех лет в Средней Азии (Каршинская степь; Шенброт, 1981). По нашим данным в Калмыкии плотность населения малого тушканчика на площадке мечения изменялась в течение 1997-2000 г с 8 до 1 особи/га. Причина падения численности – засухи, приводящие к снижению интенсивности размножения и жировых запасов перед залеганием в спячку (Утешев, 1980; Магомедов, 1987, 1989). Малый тушканчик питается семенами и зелеными частями растений. Состав диеты подвержен значительной сезонной и географической изменчивости. На большей части ареала семена служат основным кормом. Их доля по объему изменяется в пределах ареала от 41 до 85%. Участие зеленых кормов в диете (по объему) возрастает с севера на юг с 14 до 49%. На крайнем юге Средней Азии поедают семена и зеленые части растений примерно в равном соотношении. Подземные части растений и корма животного происхождения – постоянные, но незначительные по объему компоненты диеты (Шенброт и др., 1995). Значительная роль подземных частей растений в питании отмечена в Юго-западном Таджикистане (Давыдов, 1964, 1988), а преобладание насекомых в Ферганской долине Узбекистана (Павленко, Давлетшина, 1971). Сезонная изменчивость питания определяется сроками массового созревания основных кормов – семян травянистых растений. Так в Северных, Северо-западных Кызылкумах и на Устюрте весной до начала плодоношения в желудках преобладали зеленые части растений, летом доминировали семена, а осенью вновь возрастала доля зеленых кормов (Сабилаев, 1971; Бекенов, Мырзабеков, 1974). Южнее в Каршинской степи Узбекистана весной основную роль в составе кормов играли семена эфемеров, летом, напротив, возрастало значение зеленых растений-суккулентов, а осенью – снова увеличивалась доля семян (плодов суккулентов; Шенброт, 1981). Характерны два типа нор – летние постоянные и зимовочные. Собственных защитных нор, как правило, нет (Мазин, 1973; Роговин, 1983), или число их мало (одна защитная на две постоянных; Давыдов, 1988). В типичном случае летняя постоянная нора состоит из идущего от забитого земляной пробочкой входа, наклонного хода, на конце которого расположена гнездовая камера. От основного хода вверх отходит один или несколько отнорков (до 8, обычно 1-2), немного не доведенных до поверхности и служащих запасными выходами. Начальный ход, используемый при строительстве норы, как и у других видов пятипалых тушканчиков плотно забит землей. Только 25% нор не имеют запасных выходов. Общая длина ходов составляет 80-160 см (до 310 см), глубина залегания гнездовой камеры – как правило, 40-70 см. Гнезда имеются практически во всех норах самок и только в половине нор самцов. Диаметр гнездовой камеры – 10-13 см. Гнездо сооружают из размочаленных стеблей растений, чешуек луковиц, клочков шерсти, пуха перьев (Шенброт и др., 1995). Летние постоянные норы используются от нескольких суток до двух с лишним месяцев (Мазин, 1973). Зимовочные норы отличаются присутствием специальной камеры спячки и значительно большей глубиной (130-135 см в Южном Прибалхашье (Исмагилов, 1961; Мазин, 1973) и 110-160 см в Таджикистане (Давыдов, 1988). В Закавказье в связи с отсутствием зимней спячки существует только один тип нор (Аргиропуло, 1939; Алиева, 1965), причем летние постоянные норы там имеют относительно более простое строение. При строительстве норы для разрыхления грунта используют в первую очередь резцы. Тушканчик производит серию ударов резцами о грунт с частотой около 9 ударов в секунду. В результате образуется щель в грунте, и зверек откусывает комок (Гамбарян и др . , 1978). Грунт отгребают передними лапками или отодвигают мордочкой развернувшись. Затем отбрасывают синхронными ударами задних лап. В Средней Азии и Казахстане зимняя спячка продолжается от 2,5 до 4,5 месяцев в зависимости от погодных условий года. Первыми залегают взрослые самцы, затем взрослые самки и, наконец, сеголетки. На севере залегание начинается с середины октября, на юге с середины- конца ноября, пробуждение начинается на севере с середины марта, на юге с середины февраля. При кратковременных оттепелях в середине зимы на юге ареала могут пробуждаться и оставаться активными по несколько дней (Шенброт и др., 1995) В Восточном Закавказье вообще не впадает в зимнюю спячку, однако активность зимой заметно снижается (Алиева, 1965; Эйгелис, 1980). Максимальная продолжительность жизни в природе 2-2,5 года (Лобачев, Шенброт, 1977). Активность сумеречная и ночная. Весной выходят из нор в основном через 15-25 минут после захода солнца, летом через 45 мин. Весной и летом взрослые активны в течение всей ночи. Худых зверьков иногда можно встретить и за 2-3 часа до захода солнца. Весной и летом активны в течение всей ночи, осенью в основном в первой половине ночи (Сабилаев, 1971; Слудский и др., 1977; Давыдов, 1988) Характерны три типа бипедальной локомоции. Наиболее типичен асинхронный рикошетирующий прыжок, наблюдаемый при быстром беге. При этом толчок производится сначала одной, затем другой лапой (Фокин, 1978). При медленном передвижении прыжками отталкиваются задними лапами одновременно, либо передвигаются шагом, однако, последний аллюр наблюдается много реже, чем у крупных представителей Allactaga (Роговин, 1999). Максимальная скорость бега 8,3 м/сек при длине прыжка 202 см (Фокин, 1978). При преследовании спасаются бегством, совершая резкие броски из стороны в сторону, часто меняют направление движения и совершают вертикальные прыжки. В норы при преследовании забегают очень редко (Роговин, 1999). Умеренно агрессивны. По наблюдениям в природе лишь 35% контактов имеют вид направленной агрессии, в остальных случаях при встрече зверьки просто разбегаются в разные стороны (Роговин, 1983) Участки обитания сравнительно крупные. В Западном Казахстане при плотности около 2 особей/га площадь участка самцов составляла в среднем 1,06 га, у самок), 0,81 га (Роговин, 1983). В Юго-восточном Казахстане среднее удаление от постоянной норы составляло 121 м (от 32 до 296 м), т.е. средние размеры участков составляли 4,6 га (Мазин, 1973). В разреженных поселениях участки самок перекрываются лишь частично, при этом распределены более равномерно, чем участки самцов, каждый из которых перекрывает участки 1-3-х самок. В плотных поселениях участки обитания обоих полов могут полностью перекрываться (Роговин, 1983,данные Г.И. Шенброта). В течение трех месяцев В.Н. Мазин наблюдал смещение центра участка обитания на 760 м. (Мазин, 1973). В Казахстане и Средней Азии приступают к размножению сразу после выхода из спячки. В Закавказье, где спячка отсутствует начинают размножаться уже в феврале. Во всех случаях имеются два пика интенсивности размножения: весенний в апреле и осенний – в августе или сентябре. В середине лета репродуктивная активность гонад падает. В течение года большинство самок приносит по два, а часть по три выводка. При повторном весеннем размножении наблюдаются случаи беременности совмещенной с лактацией. В осеннем размножении наряду со старыми самками размножаются и рожденные весной сеголетки (Шенброт и др., 1995) Обычно в весеннем размножении участвуют все самки; лишь в годы сильных засух часть самок не размножается весной (Утешев, 1980). Весеннее размножение сильно растянуто: в первую очередь в него вовлекаются двухлетние самки, затем годовалые и лишь в конце родившиеся осенью самки предыдущего года. Интенсивность повторного раннелетнего размножения заметно ниже. В обычные годы в нем участвует до 30% всех половозрелых самок, в первую очередь самки старших возрастов. Молодые самки (осенние) повторно в это время не размножаются. В засушливые годы раннелетнее размножение отсутствует (Мазин, 1975; Павленко, 1975; Шенброт, 1980). Участие в осеннем размножении зависит от погодных и кормовых условий. В благоприятные годы в нем принимают участие практически все самки, включая рожденных весной (Бекенов, Мырзабеков, 1977; Шенброт, 1980). В выводках 1-9, обычно 3-6 детенышей. Самые крупные размеры выводков наблюдаются во время весеннего размножения, меньшие во время осеннего и самые маленькие во время раннелетнего (повторного за весенним) размножения (Мазин, 1975; Бекенов, Мырзабеков, 1977; Шенброт, 1980). Беременность длится, судя по косвенным данным, 20-21 день (Салихбаев, 1973).Эмбриональная смертность низкая (1-4%; Мазин, 1975; Давыдов, 1988). Масса тела новорожденных 2,5-2,6 г. К самостоятельной жизни переходят в возрасте 35-40 дней. В возрасте 25 дней – еще слепые (Исмагилов, 1948, 1961; Бекенов, Мырзабеков, 1977). Ко времени выхода из норы масса тела молодых составляет 21-26 г (в среднем 25,6 г). Молодые, рожденные весной, достигают половой зрелости в возрасте 2,5-3,5 месяцев, а рожденные осенью – в возрасте 9 месяцев (Шенброт и др., 1995). Соотношение полов среди новорожденных близко к 1. Такое же соотношение сохраняется, как правило, и среди взрослых. Известные отклонения могут быть объяснены различной сезонной активностью полов (Шенброт и др., 1995). Линька взрослых в Средней Азии и Казахстане протекает после окончания первой волны размножения с начала июня до сентября (Сабилаев, 1971) По данным Г.И. Шенброта пик интенсивности линьки взрослых приходится на июль (Шенброт и др., 1995). Алиева Ш.Б. 1965. Материалы по фауне и экологии грызунов Нахичеванской АССР // Тр. ин-та зоол. АН Азерб. ССР. Т. 25. С. 133-164. Алтухов А.А., Трощенко Б.В., Лаврентьев А.Ф.1969. Некоторые вопросы экологии и экспериментальной чумы у тушканчиков Зауралья // Пробл. особоопасн. инфекций. Вып. 6. С. 45-47. Андрушко А.М. 1939. Деятельность грызунов на сухих пастбищах Средней Азии. Л.: Изд. ЛГУ,. 154 с. Аргиропуло А.И. 1939. К распространению и экологии некоторых млекопитающих Армении // Зоол. сб. Т. 1. Тр. биол. ин-та Арм. фил. АН СССР. Вып. 3.. С. 27-66. Бекенов А., Мырзабеков Ж. 1974. Особенности питания тушканчиков Северцова и малого на пустынных пастбищах северного Кызылкума // Вестн. с./х. наук Казахстана. Т. 12. С. 58-62. Бекенов А., Мырзабеков Ж. 1977. Размножение малого тушканчика (Allactaga elater ) в северном Кызылкуме и на Устюрте // Зоол. журн. Т. 56, вып. 5. С. 769-778. Бибиков Д.И., Стогов И.И.1963. Материалы по млекопитающим Чингизтау // Бюлл. МОИП, отд. биол. Т. 68. вып. 4. С. 14-23. Виноградов Б.С. 1937. Фауна СССР. Млекопитающие. Т. 3, вып. 4. Тушканчики. М.-Л.: Изд. АН СССР. 197 с. Виноградов Б.С., Аргиропуло А.И. 1941. Фауна СССР. Млекопитающие. Определитель грызунов. М.-Л.: Изд-во АН СССР. 243 с. Воронцов Н.Н., Малыгина Н.А., Раджабли С.И. 1971. Хромосомы тушканчиков (Rodentia, Dipodidae) // Зоол. журн. Т. 50, вып. 12. С. 1853-1860. Гамбарян П.П., Зубцова Г.Е., Зыкова Л.Ю., Крастс И.В. 1978. Особый тип разрыхления земли резцами пятипалых тушканчиков//Зоол. журн. Т. 57, вып. 5. С. 806-809. Давыдов Г.С. 1964.Грызуны северного Таджикистана. Душанбе: Изд. АН Тадж. ССР. 272 с. Давыдов Г.С. 1988. Фауна Таджикской ССР. Том ХХ, часть 3. Млекопитающие (грызуны). Душанбе: Дониш. 315 с. Ефимов В.И., Зархидзе В.А. 1964. Материалы по фауне и экологии грызунов пустынь северного Прикарабугазья // Изв. АН Туркм. ССР, сер. биол. н. Вып. 3. С. 51-55 Исмагилов М.И. 1948. К экологии зайца-русака, малого тушканчика и серого хомячка на о. Барса-Кельмес // Изв. АН Каз. ССР, Т. 63, сер. зоол., вып. 8. С. 79-93. Исмагилов М.И. 1961. Экология ландшафтных грызунов Бетпак-Далы и южного Прибалхашья. Алма-Ата: Изд. АН Каз. ССР. 368 с. Кузнецов А.И., Кузнецова М.А., Райымбеков Д.С. 1985. Тушканчики Тау-Кумов и степи Джусандала // Тушканчики фауны СССР. М.: Всесоюз. териол. о-во. С. 46-47. Лобачев В.С., Сабилаев А.С., Шенброт Г.И. 1976а. Распределение по биотопам и численность тушканчиков Приаралья // Фауна и экология грызунов, вып. 13. М.: Изд-во МГУ. С. 13-40. Лобачев В.С., Шенброт Г.И. 1977Определение возраста и возрастная структура популяции тушканчиков//Бюлл. МОИП, отд. биол. Т. 82, вып. 2. С. 18-23. Магомедов М.-Р.Д. 1987. Половая структура популяции некоторых зимоспящих видов грызунов // Экология. Вып. 2. С. 13-20. Магомедов М.-Р.Д. 1989. Зависимость питания и состояния популяций тушканчиков от динамики кормовых ресурсов // Журн. общ. биол. Т. 50, вып. 6. С. 824-835. Мазин В.Н. 1973. Тушканчики юго-востока Казахстана и их хозяйственное значение: Автореф. дисс. канд. биол. наук. Алма-Ата. 26 C. Мазин В.Н. 1975. Материалы по размножению тушканчиков рода Allactaga на юго-востоке Казахстана // Экология, Вып. 4. С. 53-60. Миронов Н.П., Карпузиди К.С., Клименко И.З., Колесников И.М., Лисицын А.А., Нельзина Е.Н., Ширанович П.И., Ширяев Д.Т., Яковлев М.Г. 1965. Источники и переносчики чумы и туляремии. М.: Медицина, 195 с. Нургельдыев О.Н. 1960. Материалы по фауне и экологии млекопитающих трассы Каракумского канала первой очереди. Ашхабад: Изд-во АН Туркм. ССР. 287 с. Огнев С.И. 1948. Звери СССР и прилежащих стран. Т. 6. М.-Л.: Изд-во АН СССР. 559 c. Павленко Т.А. 1975. Размножение малого тушканчика в Ферганской долине // Экология, Вып. 6. С. 99-101. Павленко Т.А., Давлетшина А.Г. 1971. Питание малого тушканчика в Ферганской долине // Экология, Вып. 1. С. 91-93 Прокопов К.П. 1985. О тушканчиках Зайсанской котловины // Тушканчики фауны СССР. М: Всесоюз. териол. о-во. С. 60-62. Рабинович Б.К., Ахундов М.А. 1975. Материалы о численности и подвижности малого тушканчика в Бозчели // Пробл. особо опасных инфекций. Саратов, Вып. 3-4. С. 33-36. Роговин К.А.1983. Структура популяций и поведение тушканчиков: Автореф. дисс. канд. биол. наук. М.: ИЭМЭЖ АН СССР. 23 с. Роговин К.А. 1999. К вопросу о происхождении бипедальной локомоции у грызунов (Экологические корреляты двуногого бега тушканчикообразных) // Зоологический журнал 78 (2): 228-239 Сабилаев А.С. 1967 Размещение и численность тушканчиков в Каракалпакии и прилежащей территории//Экология млекопитающих и птиц. М.: Наука. С. 60-66. Сабилаев А.С. 1971. К экологии малого тушканчика ( Allactaga elater ; Mammalia, Dipodidae) на Устюрте и в северо-западных Кызылкумах // Науч. докл. высш. школы. Биол. н. Вып. 2. С. 15-21. Салихбаев И.К. 1973. Внутриутробное развитие малого тушканчика // Узб. биол. журн. Вып. 3. С. 44-46. Слудский А.А., Бекенов А., Борисенко В.А., Грачев Ю.А., Исмагилов М.И., Капитонов В.И., Страутман Е.И., Федосенко А.К., Шубин И.Г., 1977. Млекопитающие Казахстана. Т. 1, ч. 2. Алма-Ата: Наука. 536 с. Сухов В.П. 1970. Позднеплиоценовые мелкие млекопитающие Аккулаевского местонахождения в Башкирии. М.: Наука. 94 с. Утешев Х. 1980. Влияние засухи на размножение некоторых видов тушканчиков на Устюрте // Вестн. Каракалп. фил. АН Уз. ССР. Вып. 2. С. 51-53. Фокин И.М. 1978. Локомоция и морфология органов движения тушканчиков. Л.: Наука. 119 с. Ходашева К.С. 1953. Жизненные формы грызунов равнинного Казахстана и некоторые закономерности их географического распространения // Тр. ин-та географии АН СССР. Т. 54. С. 33-194. Шенброт Г.И. 1980. Сравнительная экология тушканчиков пустынь Турана. Автореф. дисс. канд. биол. наук. М.: МГУ. 22 с. Шенброт Г.И. 1981. Разделение ресурсов между совместно обитающими видами тушканчиков (Rodentia, Dipodidae) в Каршинской степи // Зоол. журн. Т. 60, вып. 4. С. 557-567. Шенброт Г.И. 1984. Строение коренных зубов и филогения пятипалых тушканчиков подсемейства Allactaginae (Rodentia, Dipodidae) // Сб. тр. зоол. музея МГУ. Т. 22. С. 61-92. Шенброт Г.И. 1991 Ревизия подвидовой систематики пятипалых тушканчиков рода Allactaga фауны СССР // Тр. зоол. ин-та АН СССР,. Т. 243. С.42-58. Шенброт Г.И., Соколов В.Е., Гептнер В.Г., Ковальская Ю.Г., 1995. Млекопитающие России и сопредельных регионов. Тушканчикообразные. М.: Наука. С 1-573. Эйгелис Ю.К. 1980. Грызуны Восточного Закавказья и проблема оздоровления местных очагов чумы //Саратов: Изд-во Саратовск. ун-та. 262с.
|   |
||||||||||||||||||||||||
| © 2000-2026 гг. Центр охраны дикой природы. Все права защищены |